![]()
Home
Mission
Overview of Project
Project Staff
Sponsors
Achievements
Checking, Illustrations
Upcoming Activities
Id and Species Lists
Protea Information
Protea Gallery
Growing Proteas
Interim Dist. Maps
Publications
Afrikaanse Inligting
![]()
Hybrids Relationships within Proteas
| SASOL Proteas puts the genera in groupings based on similarity. The hope is that these are ‘natural groupings’ – species that are closely related. This not only makes the species easy to identify, but also tells us a lot about their evolution. There are two other methods of assessing natural groupings: |
| Look at their DNA distances – in essence compare how much their DNA has changed and therefore how long ago they split from their sister species. There are two important assumptions in this method – that DNA mutations are constant within a gene over the timespan of the species and lineages considered, and that these mutations stay constant during the speciation event. Tim Barraclough and Gail Reeves are doing this for Sugarbushes, using specimens collected by us, and we hope to report on their preliminary and final results in the next few issues. Nigel Barker is doing the same for the Conebushes.
| Look at how much they interbreed. Species that interbreed most are more likely to be closely related than those that do not. There are problems with this assumption. For instance, where closely related species share distribution ranges, they will have evolved barriers to interbreeding (or else have fused into a single species) Conversely, less closely related species, that are separated geographically, have no need to evolve isolating mechanisms. Also, there is a host of potential mechanisms by which isolation can be effected, some of which are better at preventing hybridization than others. These mechanisms vary in effectiveness. And mutations can disable some of these systems allowing less-related species (or populations within a species – or even freak plants) to interbreed with other species.
The Protea Atlas Project has a large collection of data on hybrids found in nature. We can use these data to interpret relationships between species, provided that we remember the limitations of the data.
Firstly, we only have natural data. These are not controlled experiments, and thus we do not have data on species that do not share distribution ranges. Many closely-related species occur in mutually exclusive areas, and in these cases we will not have any data on hybridization (correctly – as it does not occur), but the species may hybridize heavily if they are planted together. Thus the species will appear to be unrelated from Protea Atlas data, but experiments will give a different conclusion.
Secondly, different species may hybridize to different degrees. We have data on both numbers of localities in which hybrids have been seen, and on the numbers of hybrids seen in each case. Some species’ pairs may produce large numbers of hybrids at only a few locations (and none at others), and others may produce only a very few plants in most areas where they co-occur. These are two different types of data, which can be teased out in controlled experiments, but that cannot be strictly compared using atlas data.
Thirdly, some hybrids are easy to spot. Or it may be impossible to identify a hybrid because it will merely resemble one of the parent species. Occasionally hybrids may back-cross with one of both of the parents giving rise to intermediates which may resemble them, and thus atlassers will underestimate or not even notice the hybrids. We have assumed that atlassers – having both parents and the hybrids in hand, are far better at identifying hybrids than even the experts, who are usually asked to prognosticate on a hybrid species without the parent specimens. In variable species, hybrids may be mis-classified by experts if specimens sent in resemble populations of one of the parent species from some other locality. Thus, for variable species field intuition is far more reliable than herbarium comparisons. However, variable species are likely to have their hybrids over-looked – even in the field – than more constant species.
So, bearing all the above in mind, what do our data tell us?
Firstly, there are no intergeneric hybrids. The only known exception is Diastella thymelaeoides X Leucospermum oleifolium known from a single plant at Sunny Seas (Betty’s Bay). What is curious about this record, is that Di thym no longer occurs at Sunny Seas according to Protea Atlas data – the Diastella that occurs there now is Di fraterna. It is generally accepted that these two genera are very closely related, so a single plant will not upset our ideas.
Among the Sugarbushes, it is obvious that the Spoonbract and Bearded groups and the Wagon Tree Pr niti, with their large distribution ranges, account for most of the hybridization recorded. Pr burchellii stands out for its extensive hybridization with Pr laurifolia. Pr longifolia and Pr laurifolia have produced a large variety of crosses. Conspicuously absent from the list is the King Pr cynaroides, which one would expect to have at least a few hybrids based on its distribution, suggesting that it is distinct to the rest of the genus. Also there are no hybrids between Rodent and other Sugarbushes, making the same point. There are no outright surprises – most groups behaving as one would expect based on SASOL.
Among the Conebushes, it is the Sunshine and Deltaseed species that have most of the hybrids, these again being the groups with widest distribution ranges within species. Specifically, Ld coniferum, Ld gandogeri, Ld laureolum and Ld xanthoconus hybridize either with many species or produce large hybrid populations relative to more widespread species. Conspicuous here are the large number of known hybrids which have not been atlassed – some of these deficiencies in our data are due to local species extinctions (e.g. Ld levi and Ld flor). Again, given the distribution ranges, there are no surprizes compared to SASOL groupings. However, more hybrid data to link the isolated species would be appreciated.
By contrast, the number of hybrids recorded for the Pincushions (and allies) is very low. Only among the Sandveld Pincushions is hybridization rampant. Specifically, Ls hypophyllocarpodendron and Ls parile and also Ls rodolentum and Ls tomentosum appear to be having an affair. However, few Pincushions overlap in distribution ranges: Ls cuneiforme is conspicuous by its absence from the list. Other species pairs where hybrids are not as frequent as one might expect are Ls prostratus vs Ls penduncalatum, Ls tottum vs Ls vestitum, and Ls cordifolium vs Ls patersonii. As a rule, within the large and small flowered division of the genus, most Pincushions occur on their own, and so hybridization should not be expected. {But are stands of Pincushions planted for flower picking producing hybrids?} Hybrids amongst the Widetubed Pincushions are notoriously hard to distinguish (identifying the species is bad enough), and are probably under-recorded. Pincushions show their natural groupings better than the other genera.
Data for the remaining genera are sparse, and suggest that they behave much like the Pincushions. No hybrids are known within the Spoons, Powderpuffs, Silkystars or Vexators. We have data (49 records) for six crosses in Spiderheads, but only the Se glomerata X Se villosa cross is frequently noted (42 records). One would expect hybrids amongst the Sceptres, but we have none.
The only other genus with species with wide distribution ranges is the Featherbushes, and we have 3 records (of frequent abundance) for Au cancellata X Au umbellata.
Thus the Protea Atlas Project data suggest that the groupings in SASOL are not unreasonable. At least, given the limitations of the data, there are no major problems! Even though that is a pity (it would be nice to rattle a few cages), it is comforting to know that we seem to have the close relationships sewn up. Unfortunately, we do not have sufficient data to suggest groupings of the groups in the genera. But then that is best tackled with the DNA work.
Relationships within the Sugarbush - Protea genus
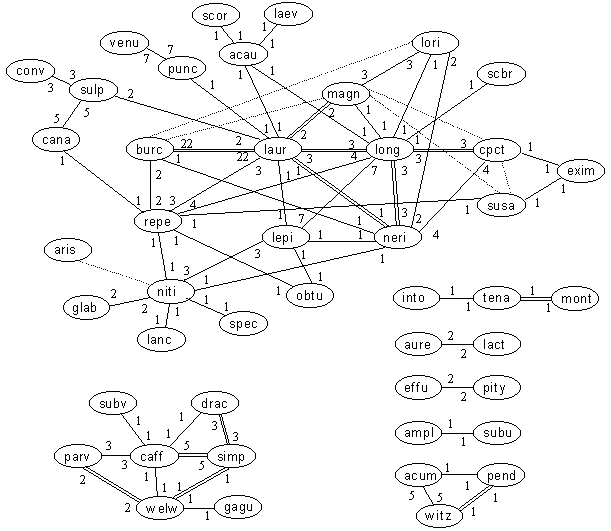
Key: Numerals are the number of records in our database. Dotted lines are literature records only. Bold lines are those for which numbers exceed 100 plants in at least one locality (ie. code "C"). Double lines are those for which numbers exceed 9 plants in at least one locality (ie. code "F").
Relationships within the Conebush - Leucadendron genus
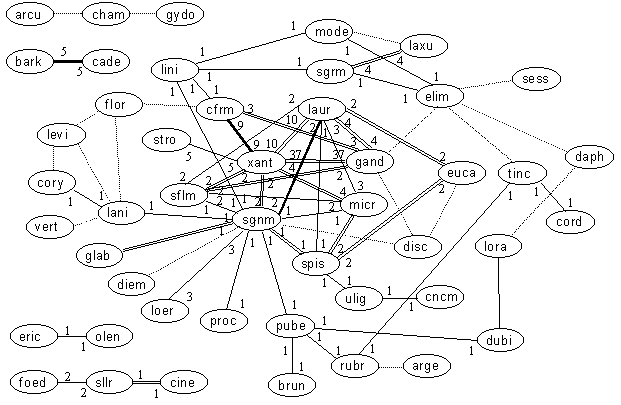
Key: Numerals are the number of records in our database. Dotted lines are literature records only. Bold lines are those for which numbers exceed 100 plants in at least one locality (ie. code "C"). Double lines are those for which numbers exceed 9 plants in at least one locality (ie. code "F").
Relationships within the Pincushion - Leucospermum genus
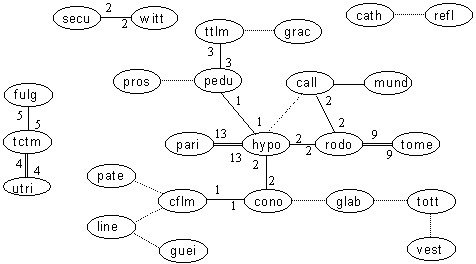
Key: Numerals are the number of records in our database. Dotted lines are literature records only. Bold lines are those for which numbers exceed 100 plants in at least one locality (ie. code "C"). Double lines are those for which numbers exceed 9 plants in at least one locality (ie. code "F").
Back Protea Ecology